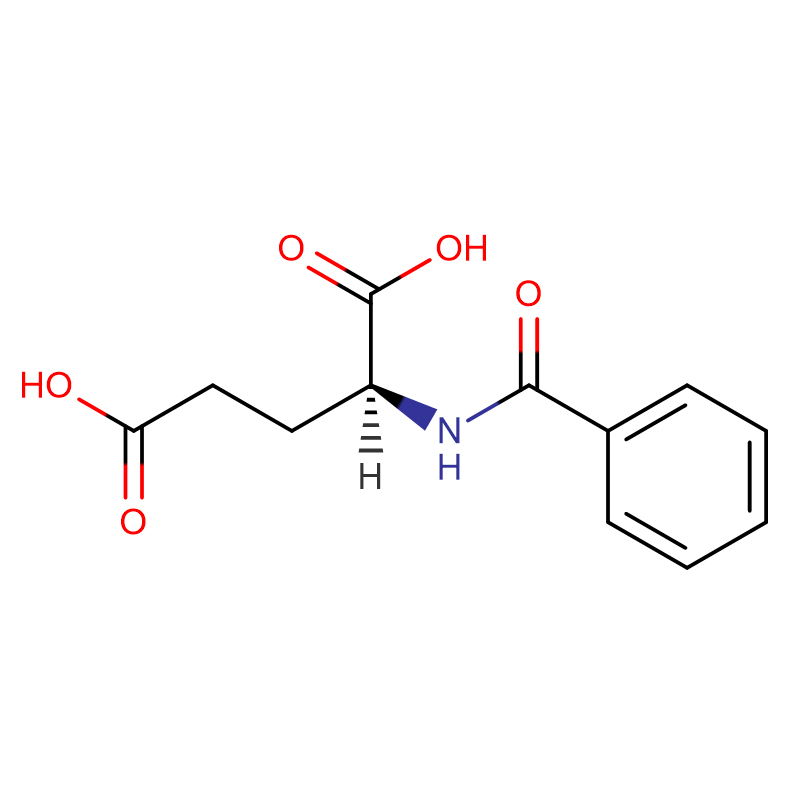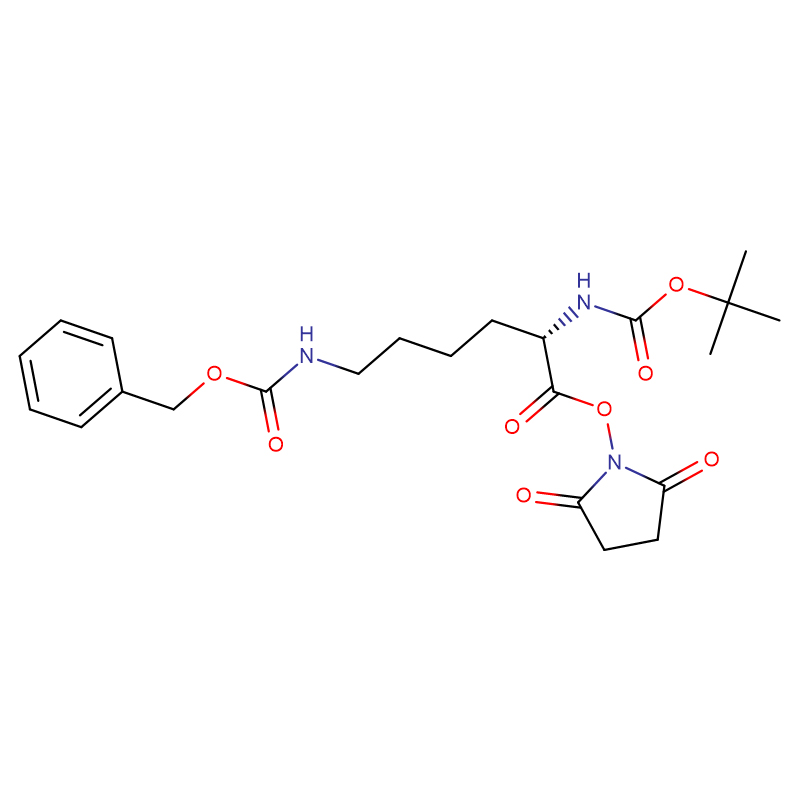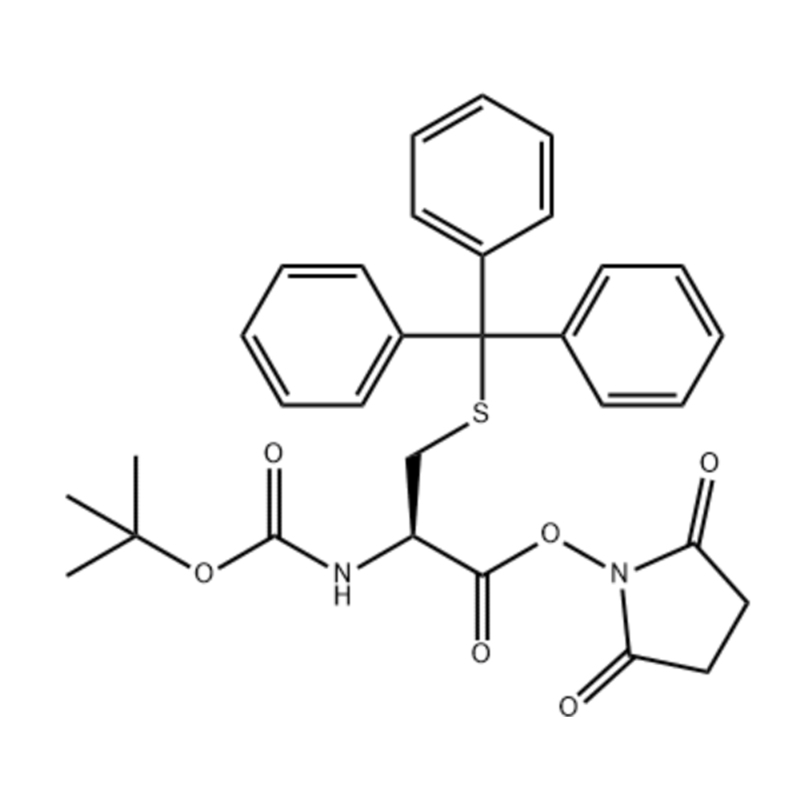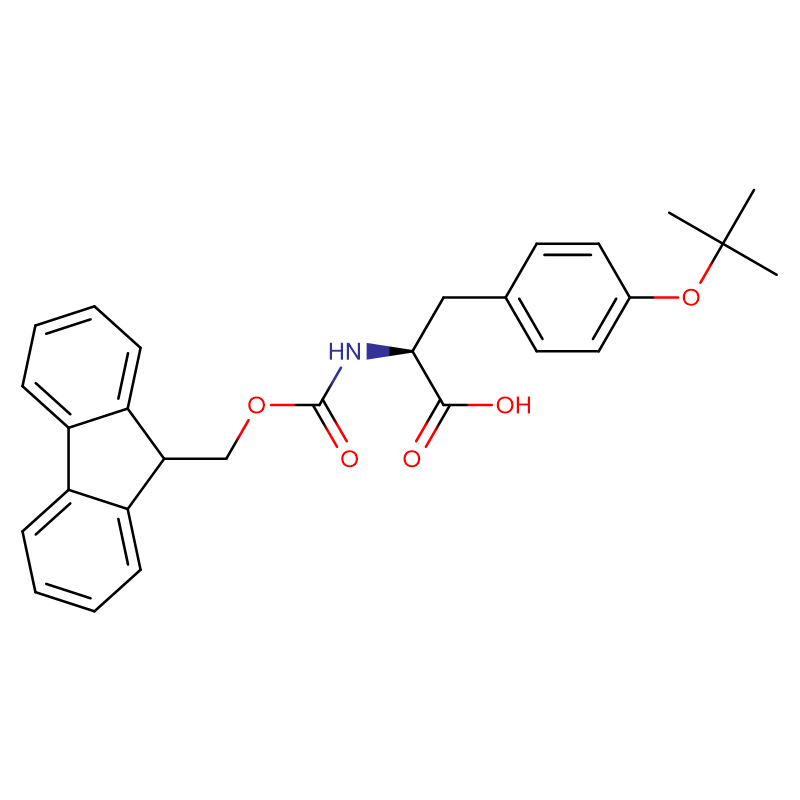
L-Methionine Cas: 63-68-3 99% ម្សៅគ្រីស្តាល់ពណ៌ស
| លេខកាតាឡុក | XD90287 |
| ឈ្មោះផលិតផល | L-Methionine |
| CAS | ៦៣-៦៨-៣ |
| រូបមន្តម៉ូលេគុល | C5H11NO2S |
| ទម្ងន់ម៉ូលេគុល | ១៤៩.២១១៣៤ |
| ព័ត៌មានលម្អិតនៃការផ្ទុក | បរិយាកាស |
| លេខកូដពន្ធដែលចុះសម្រុងគ្នា។ | ២៩៣០៤០១០ |
ការបញ្ជាក់ផលិតផល
| ការវិភាគ | 98% - 101.5% |
| រូបរាង | គ្រីស្តាល់ពណ៌សឬម្សៅគ្រីស្តាល់ |
| សេចក្តីសន្និដ្ឋាន | |
| ការកំណត់អត្តសញ្ញាណ | បំពេញតាមតម្រូវការ |
| pH | ៥.៦ - ៦.១ |
| ការបាត់បង់លើការសម្ងួត | ≤ 0.3% |
| ស៊ុលហ្វាត (SO4) | ≤ 0.03% |
| ជាតិដែក | ≤ 0.003% |
| សំណល់នៅលើភ្លើង | ≤ 0.4% |
| ក្លរ | ≤ 0.05% |
| ដែកធ្ងន់ | ≤ 0.0015% |
| ភាពបរិសុទ្ធ Chromotograh | រកមិនឃើញលើសពី 2.0% នៃសារធាតុមិនបរិសុទ្ធសរុប |
| ការបង្វិលជាក់លាក់ [ α ] D 2 5 | +22.4º ~ +24.7º |
ការសិក្សាពីមុនរបស់យើងបានបង្ហាញពីតួនាទីសំខាន់របស់វាទាំងក្នុងដំណាក់កាលដំបូង និងដំណាក់កាលចុងក្រោយនៃការឆ្លងមេរោគ KSHV ។នៅក្នុងការសិក្សានេះ យើងបានស្ទង់មតិអន្តរកម្មនៃ ORF45 ដោយប្រើបន្ទះនៃអង្គបដិប្រាណ monoclonal ។បន្ថែមពីលើប្រូតេអ៊ីន kinase គ្រប់គ្រងក្រៅកោសិកា (ERK) និង p90 ribosomal S6 kinase (RSK) ដែលបានកំណត់ពីមុន យើងបានរកឃើញប្រូតេអ៊ីន copurified ផ្សេងទៀត រួមទាំងប្រូតេអ៊ីនលេចធ្លោនៃ ∼38 kDa និង ∼130 kDa ។Mass spectrometry បានបង្ហាញថាប្រូតេអ៊ីន 38-kDa គឺជាមេរោគ ORF33 ហើយប្រូតេអ៊ីន 130-kDa គឺជាកោសិកា USP7 (ubiquitin-specific protease 7) ។យើងបានគូសផែនទីនៃដែនភ្ជាប់ ORF33 ទៅនឹងអាស៊ីតអាមីណូ carboxyl-terminal 19 (aa) ដែលត្រូវបានរក្សាទុកយ៉ាងខ្លាំងនៃ ORF45 និងដែនភ្ជាប់ USP7 ទៅនឹងគំនូរឯកភាពដែលបានរាយការណ៍នៅក្នុងតំបន់កណ្តាលនៃ ORF45 ។ដោយប្រើស្នាមប្រឡាក់ immunofluorescence យើងបានសង្កេតឃើញ colocalization នៃ ORF45 ជាមួយ ORF33 ឬ USP7 ទាំងនៅក្រោមលក្ខខណ្ឌឆ្លង និងនៅក្នុងកោសិកាដែលឆ្លង KSHV ។លើសពីនេះទៅទៀត យើងបានកត់សម្គាល់ការផ្លាស់ប្តូរទីតាំងដែលពឹងផ្អែកលើ ORF45 នៃផ្នែកមួយនៃ ORF33/USP7 ពីស្នូលទៅ t he cytoplasm ។យើងបានរកឃើញថា ORF45 បណ្តាលឱ្យមានការកើនឡើងនៃការប្រមូលផ្តុំប្រូតេអ៊ីន ORF33 ដែលត្រូវបានលុបចោលប្រសិនបើដែន ORF33- ឬ USP7-binding នៅក្នុង ORF45 ត្រូវបានលុប។លើសពីនេះ ការលុបស្ថានីយ carboxyl ដែលត្រូវបានអភិរក្សនៃ ORF45 នៅក្នុងហ្សែន KSHV បានកាត់បន្ថយយ៉ាងខ្លាំងនូវកម្រិតនៃប្រូតេអ៊ីន ORF33 នៅក្នុងកោសិកាដែលឆ្លងមេរោគ KSHV និងបានលុបចោលការផលិតនៃមេរោគ progeny ។សរុបមក លទ្ធផលរបស់យើងមិនត្រឹមតែបង្ហាញសមាសធាតុថ្មីនៃអន្តរកម្ម ORF45 ប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏បង្ហាញថាអន្តរកម្មក្នុងចំណោមប្រូតេអ៊ីនទាំងនេះមានសារៈសំខាន់សម្រាប់ការចម្លងមេរោគ KSHV lytic ។ Kaposi របស់ sarcoma-sociated herpesvirus (KSHV) គឺជាភ្នាក់ងារបង្កនៃជំងឺមហារីកមនុស្សមួយចំនួន។KSHV ORF45 គឺជាប្រូតេអ៊ីនពហុមុខងារដែលត្រូវបានទាមទារសម្រាប់ការចម្លង KSHV lytic ប៉ុន្តែយន្តការពិតប្រាកដដែល ORF45 អនុវត្តមុខងារសំខាន់ៗរបស់វាមិនច្បាស់លាស់។ការសិក្សាពីមុនរបស់យើងបានបង្ហាញថាប្រូតេអ៊ីន ORF45 ទាំងអស់នៅក្នុងកោសិកាមាននៅក្នុងស្មុគស្មាញដែលមានទម្ងន់ម៉ូលេគុលខ្ពស់។ដូច្នេះហើយ យើងបានព្យាយាមកំណត់លក្ខណៈអន្តរកម្មនៃ ORF45 ដើម្បីផ្តល់នូវការយល់ដឹងអំពីតួនាទីរបស់វាកំឡុងពេលចម្លង lytic ។ដោយប្រើបន្ទះនៃអង្គបដិប្រាណ monoclonal យើងបានស្ទង់មតិអន្តរកម្ម ORF45 នៅក្នុងកោសិកាដែលឆ្លង KSHV ។យើងបានកំណត់អត្តសញ្ញាណដៃគូចងថ្មីពីរនៃ ORF45៖ ប្រូតេអ៊ីនមេរោគ ORF33 និងកោសិកា ubiquitin-specific protease 7 (USP7)។យើងបង្ហាញបន្ថែមទៀតថាអន្តរកម្មរវាង ORF45 និង ORF33 គឺមានសារៈសំខាន់សម្រាប់ការផលិតប្រកបដោយប្រសិទ្ធភាពនៃភាគល្អិតមេរោគ KSHV ដោយបង្ហាញថាការជ្រៀតជ្រែកដែលមានគោលដៅជាមួយអន្តរកម្មនេះអាចតំណាងឱ្យយុទ្ធសាស្រ្តប្រលោមលោកដើម្បីទប់ស្កាត់ការចម្លង KSHV lytic ។